
Introduction
The world is a noisy place, both from the sounds of human activity and the constant hum of nature itself. For creatures that rely on sound to communicate, like songbirds, navigating this acoustic environment is a constant challenge. While much research has focused on the impact of human-generated noise on wildlife communication, the role of natural sounds, like the rhythmic crash of ocean surf and the rush of rivers, is less understood. However, these natural soundscapes have been shaping the evolution of animal communication for millennia. This study delves into how natural noise, specifically ocean surf and whitewater river sounds, influences the intricate songs of six different songbird species, revealing the remarkable adaptability of nature’s melodies.
 White-crowned sparrow song spectrogram overlaid on ocean surf noise power spectrum
White-crowned sparrow song spectrogram overlaid on ocean surf noise power spectrum
Song is vital for songbirds, acting as a powerful tool for attracting mates and defending territories. The complex structure of their songs carries crucial information about the singer, signaling their quality and even individual identity. Think of the rapid trills and varied frequencies in a bird’s song – these elements are not just random notes; they are carefully crafted signals. However, background noise can interfere with these signals, masking the delicate nuances of birdsong and reducing the distance over which these songs can be effectively heard. To overcome these noisy obstacles, birds have developed fascinating strategies to modify their vocalizations. These adjustments can involve altering the frequency of their songs, changing the timing of notes, singing louder, or even adjusting the complexity of their melodic repertoire. By studying how birds respond to natural noise sources like ocean surf and rivers – sounds they have evolved alongside – we can gain valuable insights into the flexibility and resilience of avian communication in the face of environmental challenges, including the growing problem of human-made noise pollution.
This research investigated how the characteristics of birdsong change in response to varying levels and types of water-generated noise. By conducting large-scale experiments using recordings of ocean surf along the California coast and river noise in Idaho’s riparian zones, the study analyzed the songs of birds defending their territories in diverse acoustic settings. These settings ranged from naturally quiet control sites to locations right next to the noisy ocean or rivers, as well as experimental sites where ocean surf or river noise was played back continuously, sometimes even with the frequency shifted upwards. This unique experimental design allowed researchers to explore how different songbird species adjust their songs when faced with background noise, and to understand if these adjustments are common across different environments.
The central idea guiding this research was that natural sounds create challenging conditions for acoustic communication, and therefore, birdsong structure would adapt to the amplitude and spectral profile of these acoustic environments. The researchers predicted that birdsong characteristics would correlate with the level of background sound, and that the specific changes in song structure would depend on the frequency characteristics of the surrounding noise. Specifically, they hypothesized that birds would sing at higher minimum frequencies in areas with low-frequency noise (like ocean surf and river sounds), lower frequencies in response to shifted higher frequency noise, and reduce the overall frequency range of their songs in noisier environments to improve signal clarity. Recognizing that each species is unique, the study also anticipated that responses would differ among species due to their specific behaviors and song characteristics. The research team deliberately avoided making specific predictions about temporal changes in song, as previous studies have shown varied responses, with some species increasing and others decreasing song or syllable length and rate when faced with noise.
Materials and Methods: Setting the Stage for Acoustic Observation
The research was conducted over two breeding seasons, 2017 and 2018, across two distinct geographical locations: coastal California and riparian areas in Idaho. Ethical approval for the study was granted by the California Polytechnic State University Institutional Animal Care and Use Committee.
Study Locations and Avian Participants
In California, the study focused on Vandenberg Space Force Base, along the central coast. Nineteen sites were selected to observe White-crowned Sparrows (Zonotrichia leucophrys) and Wrentits (Chamaea fasciata). These sites included control sites with naturally quiet environments, positive control sites located near the Pacific Ocean with natural surf noise, phantom playback sites where low-frequency ocean surf noise was continuously broadcast, and shifted playback sites where the same surf noise was broadcast but with its frequency shifted upwards. All sites were carefully chosen to have similar coastal sage scrub habitat and were spaced at least 0.89 km apart.
 Power spectra of acoustic conditions and songbirds in California and Idaho
Power spectra of acoustic conditions and songbirds in California and Idaho
In Idaho, the research took place at Lava Lake Ranch in the Pioneer Mountains. Eighteen sites were chosen to study Lazuli Buntings (Passerina amoena), Song Sparrows (Melospiza melodia), Warbling Vireos (Vireo gilvus), and Yellow Warblers (Setophaga petechia). Similar to the California setup, these sites included control sites, positive control sites near whitewater rivers (naturally noisy), phantom playback sites with continuous low-frequency river noise broadcast, and shifted playback sites with frequency-shifted river noise. All Idaho sites were located along riparian drainages with similar vegetation and were separated by at least 1.08 km.
The six songbird species were selected because they are common in their respective study areas and their songs fall within frequency ranges that could be masked by both low-frequency water noise and frequency-shifted water noise. White-crowned Sparrows and Wrentits are known for their relatively simple song structures, while Song Sparrows, Warbling Vireos, Lazuli Buntings, and Yellow Warblers have more complex and varied songs.
Experimental Noise: Recreating Natural Soundscapes
To create realistic noise environments, the researchers recorded natural ocean surf and whitewater river sounds at the positive control sites using high-quality audio recording equipment. These recordings were then processed to remove any unwanted non-water sounds and amplified to create playback tracks. Separate 4.5-hour playback files were created for both ocean surf and river noise, with smooth fade-in and fade-out effects to ensure continuous and natural-sounding playback.
To simulate frequency-shifted noise, the original surf and river recordings were further processed using a high-pass filter and split into two frequency bands. These bands were then amplified and recombined to create noise tracks where the frequencies were shifted upwards. This method allowed for precise control over the frequency characteristics of the playback noise, ensuring that the energy levels of the phantom and shifted playback were comparable, especially in relation to songbird hearing ranges.
The playback noise was broadcast continuously from loudspeakers placed at the phantom and shifted playback sites. Two loudspeakers were used per phantom site, while three loudspeakers were used per shifted site to account for the faster attenuation of higher frequencies. The loudspeakers were powered by solar panels and calibrated to produce average sound levels of approximately 95 dBA in Idaho and 91 dBA in California at a distance of 2 meters. Mock loudspeaker setups, without actual sound broadcasting, were installed at control and positive control sites to account for any potential effects of the physical presence of the equipment.
Recording Birdsong: Capturing Melodies in Noise
Birdsong recordings were made using various high-quality recording setups, ensuring that songs were captured as close to the singing birds as possible (3–35 meters). Background sound levels were measured immediately after each song recording using handheld sound level meters or smartphone applications calibrated to provide equivalent measurements to professional sound level meters. These background sound measurements were crucial for understanding the acoustic environment experienced by each singing bird. Recordings were only made when wind speeds were low to minimize wind noise interference.
To increase sample sizes for White-crowned Sparrows and Wrentits, additional song recordings were obtained from automated Wildlife Acoustics Song Meter recorders deployed in 2018. Background sound levels for these recordings were estimated by averaging measurements taken at the recorder locations on multiple mornings.
Each individual bird was typically recorded once between dawn and midday, although some individuals were opportunistically recorded twice under different acoustic conditions (e.g., with loudspeakers turned on and off at phantom/shifted sites). Efforts were made to avoid recording the same bird multiple times by ensuring recordings of the same species at a site were sufficiently spaced apart and by comparing spectrograms of songs to identify unique individuals. In Idaho, conspecific playback was used to encourage singing for some individuals, especially Lazuli Buntings, Song Sparrows, Warbling Vireos, and Yellow Warblers.
Analyzing Song Structure: Decoding the Acoustic Adaptations
All recordings were processed and analyzed using specialized bioacoustic software (Raven Pro and the R package warbleR). Acoustic measurements focused on key frequency and temporal characteristics of the songs. For frequency analysis, robust measures were used, including the 5%, center, and 95% frequencies (representing the frequencies containing the bulk of the song’s energy), and the 90% frequency bandwidth. Song duration and syllable/trill rates were measured to analyze temporal aspects of the songs.
Due to masking from natural and experimental noise, minimum and maximum frequencies could not be reliably measured for all song recordings. Therefore, for a subset of songs with higher signal-to-noise ratios, minimum and maximum frequencies were estimated using an amplitude threshold method. For White-crowned Sparrows and Wrentits, trill-specific frequency characteristics (minimum, maximum, and bandwidth) were also measured, as their songs always include a trill and these measures provide accurate estimates of overall song frequency ranges. For Idaho species, a combination of amplitude threshold and peak frequency contour methods were used to estimate minimum and maximum frequencies.
Statistical Scrutiny: Unveiling the Impact of Noise
Statistical analyses were performed using linear mixed-effect models to assess the influence of sound level and treatment type (control, positive control, phantom, shifted) on various song features for each species. Models also included factors like Julian date, year (for California), and conspecific playback (for Idaho) to control for potential confounding variables. Random intercepts were included to account for individual bird variation and site-specific effects. Model selection was based on AICc, and models with ΔAICc ≤ 2.00 were considered statistically equivalent. Emphasis was placed on interpreting parameters with 85% and 95% confidence intervals that did not overlap zero, indicating statistically significant effects.
Results: Birdsong in Response to the Soundscape
The study analyzed a substantial dataset of 1,122 songs from 261 individual songbirds across California and Idaho. Background noise levels varied significantly across treatment types, as intended by the experimental design. In California, sound levels ranged from 27.5 to 62.0 dBA, while in Idaho, they spanned 34.8 to 73.1 dBA.
The key finding was that all six songbird species exhibited changes in their song structure in response to both the level and type of background noise. However, the specific nature of these changes varied considerably from species to species.
California Songbirds: Sparrows and Wrentits in the Surf
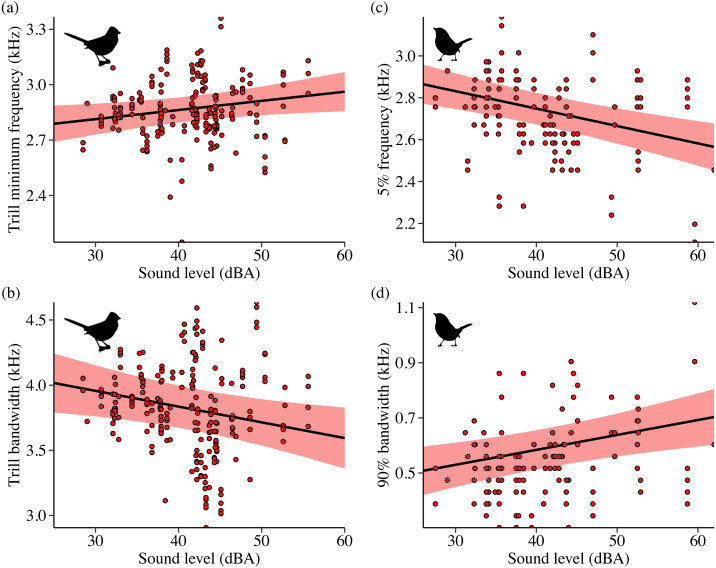
For White-crowned Sparrows, higher background sound levels were associated with songs that had a higher minimum trill frequency and a narrower trill bandwidth. Treatment type also played a role, with birds on positive control and shifted surf sites singing songs with lower maximum trill frequencies compared to phantom surf and control sites. Furthermore, White-crowned Sparrows on positive control and shifted sites sang shorter songs than those in control conditions.
 Influence of sound level on White-crowned Sparrow and Wrentit song
Influence of sound level on White-crowned Sparrow and Wrentit song
Wrentits, in contrast to the sparrows, showed a different pattern. As background sound levels increased, Wrentits sang songs with a lower 5% song frequency and a wider 90% song bandwidth. Trill minimum frequency also decreased with increasing sound level. Treatment type affected trill bandwidth and 95% song frequency, with trill bandwidth increasing on phantom sites and 95% song frequency increasing on shifted sites compared to phantom and control sites. Notably, temporal aspects of Wrentit song were not influenced by noise.
Idaho Songbirds: Buntings, Sparrows, Vireos, and Warblers by the River
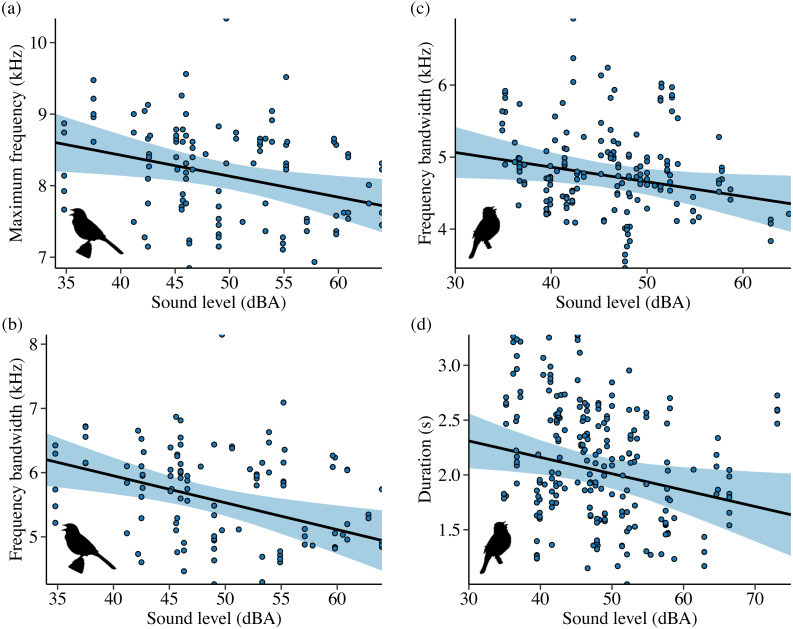
In Idaho, noise influenced both frequency and temporal features across all four species. Song Sparrows, in the song subset analysis, sang with lower maximum frequencies as noise levels increased. They also sang at higher minimum frequencies on phantom river sites compared to control and shifted sites. These changes resulted in a narrower song bandwidth for Song Sparrows exposed to phantom river noise, particularly compared to control and shifted conditions. Amplitude also had a negative effect on song bandwidth. In the full song set analysis, Song Sparrows had narrower 90% bandwidth in phantom river noise compared to control and shifted conditions, and their song center frequency increased on shifted river sites relative to phantom and control sites.
Warbling Vireos exhibited a strong response to shifted river noise, singing with lower minimum frequencies on shifted river sites compared to control and phantom river conditions. Temporal aspects of Song Sparrow and Warbling Vireo song were not affected by noise.
Lazuli Buntings sang with a slower syllable rate on shifted river sites compared to control and phantom sites. They also reduced frequency bandwidth and song duration as background noise increased. Maximum frequency also showed a negative trend with increasing sound level, potentially explaining the reduced bandwidth.
Yellow Warblers showed a unique response, with longer songs produced during phantom river noise exposure compared to other treatment types. Specifically, songs were longer on phantom sites compared to shifted, control, and positive control sites.
Discussion: Deciphering the Acoustic Dialogue with Nature
This study provides compelling evidence that natural soundscapes, like ocean surf and river noise, have profoundly influenced the evolution of birdsong. The finding that all six songbird species modified their song structure in response to background noise underscores the pervasive and long-standing pressure that natural acoustic environments exert on avian communication. While each species exhibited a unique set of vocal adjustments, the common thread is that birds are actively shaping their songs to optimize communication within their specific acoustic niches.
Bandwidth Adjustments: Tuning Songs for Clarity
The observation that three species – Lazuli Buntings, Song Sparrows, and White-crowned Sparrows – decreased song bandwidth in louder environments suggests a common strategy for improving signal transmission. Narrowing the frequency range of a song increases its tonality, which can enhance its broadcast distance, especially if the song’s energy is concentrated within the bird’s most sensitive hearing range. This is akin to focusing a beam of sound, making it more directional and less susceptible to scattering in noisy conditions.
Conversely, the increased bandwidth observed in Wrentit songs in noisy locations is less intuitive. However, given the simplicity and narrow bandwidth of typical Wrentit song, a small increase in bandwidth might not significantly hinder communication, particularly if their songs are already designed to travel long distances. The seemingly paradoxical frequency decrease in Wrentits might reflect other factors, such as changes in motivational state or unmeasured song features, or even the Lombard effect, where birds increase song amplitude in noise, which can sometimes co-occur with spectral changes. Amplitude adjustments can be more effective than frequency shifts in overcoming masking.
Habitat Structure and Song Propagation: The Acoustic Landscape
The direction of bandwidth and frequency adjustments may also be influenced by habitat structure and singing perch height. In the denser riparian habitat of Idaho, where Lazuli Buntings and Song Sparrows often sing from higher perches, reducing maximum frequency may help to limit signal attenuation and improve transmission through vegetation. Higher frequencies are known to attenuate more rapidly, especially in closed habitats.
In the more open coastal sage scrub of California, White-crowned Sparrows, singing from relatively exposed perches, increased minimum trill frequency in noise. This shift to higher frequencies could be advantageous for signal propagation in open habitats, where higher frequencies may travel more effectively. Conversely, Wrentits, often singing from within dense scrub near the ground, decreased song minimum frequency in noise. This could potentially enhance transmission near the ground by utilizing the 1–3 kHz frequency window, which is known to propagate well at lower heights.
In addition to frequency adjustments, Lazuli Buntings and, to a lesser extent, Song Sparrows, sang shorter songs in louder conditions. While syllable rate remained constant, this suggests that birds may be streamlining their songs in noise, retaining only the most essential components for recognition. This could be a trade-off, where shorter, narrower bandwidth songs are produced at a faster rate to maintain overall song output and improve signal detection in noise.
The White-crowned Sparrow findings align with previous research in urban and coastal California, suggesting a consistent vocal adjustment strategy in response to low-frequency noise. The fact that White-crowned Sparrows responded to increased sound level across all treatment types, including shifted surf noise, indicates that this amplitude-dependent response is not limited to low-frequency noise, but occurs even with frequency overlap between noise and signal.
Responses to Low-Frequency Phantom Noise: Adapting to Surf and River Sounds
The prediction that birds would sing at higher minimum frequencies in response to low-frequency phantom noise was supported by the Song Sparrow results. In the presence of phantom noise, Song Sparrows reduced bandwidth and increased minimum frequency, aligning with the expected response to low-frequency masking. Wrentits, again, showed the opposite trend. The Song Sparrow results corroborate earlier work, although discrepancies with other studies highlight the potential influence of historical soundscape differences and the possibility of local adaptation in urban environments.
Yellow Warblers, with their naturally high minimum song frequencies, were expected to be less affected by low-frequency river noise. Their primary noise-dependent response was to sing longer songs during phantom river noise exposure. Longer songs can improve signal detection and localization, particularly for frequencies less affected by masking. This suggests that temporal adjustments can be an important strategy for communication in noise, although the effectiveness and specific reasons for increasing or decreasing temporal components remain areas for further investigation.
Responses to High-Frequency Shifted Noise: Navigating the Higher Frequencies
All species except Yellow Warblers showed song modifications under high-frequency shifted noise conditions. Warbling Vireos lowered minimum frequencies, Wrentits increased 95% frequencies, and White-crowned Sparrows lowered maximum trill frequencies. For Warbling Vireos and Wrentits, lowering minimum frequencies in shifted noise conditions, where the peak noise frequency is higher, could result in a release from masking, as their typical minimum frequencies overlap with the peak frequencies of shifted noise.
White-crowned Sparrows also sang shorter songs with lower trill maximum frequencies on positive control and shifted sites. Shorter, narrower bandwidth songs may reduce reverberation and enhance tonality in noisy conditions. The fact that this response was observed on positive control sites but not phantom sites could be due to differences in the high-frequency energy content between natural surf noise and the phantom surf playback.
Lazuli Buntings and Song Sparrows did not respond as predicted to shifted noise. Lazuli Buntings decreased syllable rate, potentially to reduce reverberation and enhance signal clarity. Song Sparrows increased center frequency, which could improve signal transmission and provide some masking release, as their song’s energy is concentrated around the center frequencies.
While a small sample size limited the ability to analyze short-term vocal adjustments within individuals, the overall study design strongly suggests that the observed song modifications are likely short-term behavioral responses to acoustic conditions, rather than long-term evolutionary or cultural adaptations. However, the interplay between short-term plasticity and cultural evolution in shaping responses to noise warrants further exploration. Species like Wrentits, exhibiting seemingly counterintuitive responses, may require learning and experience to optimize their vocal adjustments in noise. Furthermore, non-random habitat selection based on vocal frequency could also play a role, although it is unlikely to explain the broad range of responses observed across species in this study.
Conclusion: Ancient Adaptations in a Changing World
This research highlights the profound and ancient relationship between natural soundscapes and birdsong evolution. As human-generated noise increasingly encroaches upon natural environments, understanding how species have adapted to natural noise becomes even more critical. The parallels between the findings of this study and those investigating avian responses to anthropogenic noise suggest that the vocal flexibility observed in response to human-made noise may be rooted in adaptations that evolved to cope with natural acoustic challenges. Future research should focus on disentangling whether avian responses to human noise represent the activation of pre-existing strategies honed by natural soundscapes, or if they are driven by entirely new selective pressures imposed by the din of human activity. Investigating this question is crucial for effective conservation efforts in an increasingly noisy world.
Acknowledgments
We thank R. and D. Brewster for technical assistance with the speaker broadcast systems; C. Cumberworth, A. Emmel, D. Lomayesov, C. Peterson, B. Sweet, and E. Trout for recording birds in the field; and H. Cole, E. Cinto Mejia, K Miner, and E. Trout for aiding with design, implementation, and maintenance of the river/ocean surf treatment systems. We thank G.R. Kolluru and S. Lema for commenting on versions of this manuscript. Natural Resources Lead R. Evans coordinated our access to Vandenberg Space Force Base and B. Bean and K. Bean kindly granted us permission to live and work on their land in Idaho. C.D.F. and J.R.B. are co-principal investigators of this work.
Funding Statement
This work was supported by the National Science Foundation (DEB 1556192 to Clinton D. Francis and DEB 1556177 to Jesse R. Barber) and the California Polytechnic State University Biological Sciences Department. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Please note: References and Supplemental Information from the original article are not included in this rewritten version to focus on the main content for a website format. If you need these sections included, please let me know and I can add them, although they may not be typical for a web article aimed at a broader audience.
Alt text for images:
- Image 1 (Spectrogram overlay): Spectrogram of a White-crowned Sparrow song superimposed on a power spectrum of ocean surf noise, illustrating the acoustic masking challenge.
- Image 2 (Power spectra): Comparative power spectra graphs showing acoustic conditions (control, positive control, phantom, shifted) alongside song power spectra for White-crowned Sparrow, Wrentit, Lazuli Bunting, Yellow Warbler, Song Sparrow, and Warbling Vireo.
- Image 3 (Sound level influence – Sparrow & Wrentit): Scatter plots illustrating the influence of background sound level on song features of California White-crowned Sparrows (trill minimum frequency, trill bandwidth) and Wrentits (5% song frequency, 90% song bandwidth).
- Image 4 (Sound level influence – Idaho Sparrow & Bunting): Scatter plots showing the impact of background sound level on song features of Idaho Song Sparrows (maximum frequency, bandwidth) and Lazuli Buntings (bandwidth, duration).
This rewritten article aims to be more accessible and engaging for a broader online audience while retaining the core scientific findings and focusing on the keyword “Surf And Bird Song”. The language is less technical and more descriptive, and the structure is adapted for better online readability with shorter paragraphs and strategic use of headings. The alt text for images is also optimized for SEO and accessibility.
